| |
|
|
UVOD
Nepobitni dokazi o ulozi endokardnog endotela u embrionalnom
razvoju srca i funkciji miokarda potvrđeni su nakon otkrića faktora
rasta neuregulina poreklom iz enokardno endotelnih ćelija. U ranoj
embrionalnoj fazi neuregulin (NRG) se sintetiše i oslobađa iz
srčanih endotelnih ćelija i deluje kao parakrini molekul koji dovodi
do aktivacije tirozin kinaze preko ErbB2 (HER-2 ili Neu) i ErbB4
(HER-4) receptora, distribuiranih u susednim kardiomiocitima [1].
Aktivacija ErbB2/ErbB4 kompleksa receptora uz pomoć neuregulina
neophodna je za trebekulaciju primitivnog sunđerastog srca [2].
Neuregulin kao faktor rasta predstavlja specifičan i bitan korak u
endokardnoj jastučastoj formaciji aktivacijom kompleksa receptora
ErbB2/ErbB4. Receptori su izraženi u prevalvularnim mezenhimnim
ćelijama koje se nalaze u susedstvu endotelnih ćelija odgovornih za
sintezu neuregulin. Kod mutanta bez ErbB3 receptora dolazi do
abnormalnog formiranja srčanih zalistaka sa posledničnom
kongestivnom srčanom insuficijencijom i smrću u uterusu [3].
NRG-ErbB signalni put, koji je važan u embrionalnom razvoju srca,
ima svoju ulogu i u adultnom srcu. Neuregulin se uglavnom sintetiše
u endokardno endotelnim ćelijama i endotelnim ćelijama miokardnih
kapilara. ErbB3 receptori su distribuirani na endotelnim ćelijama, a
ErbB2/ErbB4 kompleks na kardiomiocitima. Solubilni NRG-1
(rekombinantni humani glijalni faktor rasta 2 ili rhGGF2) izaziva
proliferaciju kardiomiocita u embrionu, povećava preživljavanje,
utiče na apoptozu kultura kardiomiocita i indukuje hipertrofičan
rast u neonatalnim i odraslim ventrikularnim kardiomiocitima. Srčana
mitogena aktivnost zavisna od delovanja 5-HT putem aktivacije 5HT2B
receptora zavisi i od intaktnog neuregulin/ErbB signalnog puta, kako
u embrionskom, tako i u odraslom srcu. Trajno prisustvo ovih
proteina u odraslom srcu ukazuje na to da je neuregulin endotelnog
porekla bitan za funkciju miokarda, što je još jedan primer
neophodne endotelno-miokardne veze za normalnu srčanu funkciju [4].
Građa i funkcija neuregulina
Neuregulini su familija faktora rasta čiji su geni identifikovani na
ekson kodu za epidermalne faktore rasta (EGF), pronađeni kod sisara,
sa delimičnom homologijom u porodici više srodnika. Neuregulin je
prvi put bio izolovan iz nervnog tkiva gde je bilo otkriveno da
promoviše proliferaciju Švanovih ćelija i dobio je naziv glijalni
faktor rasta (GGF). Glijalni faktor rasta stimuliše fosforilaciju
tirozin kinaze putem aktivacije ErbB2 receptora i deluje na mitogenu
aktivnost Švanovih ćelija [5]. U kasnijim studijama identifikovani
su slični phospho ErbB2 stimulirajući proteini, koji su bili
pronađeni kao izoforme kodirane od jednog gena. Identifikacija još
tri gena koji kodiraju slične funkcionalne izoforme dovela je do
identifikacije drugih članova neuregulin familije. Radi
jednostavnosti, mi koristimo termin neuregulini da označimo sve
izoforme za bilo koji od četiri neuregulina kodirana sa
identifikovanih gena. Gen za NRG-1 je najproučavaniji, lociran na
hromozomu 8 i kod ljudi i kod miševa. NRG-1 kodira 21 ekson i smatra
se da može da dovede do više od 31 proteinskih izoformi. N
terminalne sekvence razlikuju NRG-1 izoforme kao što su tip I, II,
III, IV, V i tip VI [6].
Šest tipova neuregulin-1 izoformi je klasifikovano na osnovu
njihovih amino-terminalnih sekvenci. Izoforma tip III sadrži
cisteinom bogate domene (CRD), kao transmembranske domene. Svih 6
tipova izoformi imaju domene slične epidermalnom faktoru rasta
(EGF). Tip I, II, IV I V imaju imunoglobulinima slične domene između
N-terminalne sekvence i EGF domena, sa ili bez regiona razdvajanja,
dok su N-terminalni regioni u tipu III i VI direktno povezani samo
sa EGF domenima. Varijante se takođe generišu i zbog veza u
C-terminalnom region. Između dva regiona je C-terminalni
transmembranski domen (TMc). Najviše neuregulin-1 izoformi su
sintetisani kao transmembranski prekursor polipeptidi (pro-NRG-1) sa
EGF domenima lociranim u ekstracelularnom region, ali u tipu III,
oba, M- i C-terminalni region su locirani unutar ćelije.
Aminoterminalni region neuregulina-1 može da uključuje signalne
peptide, domene bogate u cisteinu, imunoglobulinima slične domene,
domene slične faktorima rasta i region glikolizacije. NRG-2, - 3 i
-4 oblici imaju daleko manji broj izoformi N-terminalnih sekvenci.
NRG-2 kodira dve varijante N-terminalnih sekvenci, tip 1A i tip 1B.
U svim neuregulinima je prisutan epidermalnom faktoru rasta sličan
domen, koji posreduje receptorsku vezu i može biti klasifikovan
zasnovano na C-terminalnim sekvencama EGF sličnim domenima, koji
variraju između α i β izoforma i svaki može biti prisutan u
odvojenoj varijanti. C-terminal za EGF domen je jukstamembranski
region, koji se ponaša kao mesto proteolitičkog cepanja. C-terminal
jukstamembranskog regiona je transmembranski domen, praćen sa a-, b-
ili c- tipom citoplazmatičnih nastavaka. Posebnost građe i
funkcionalne veze svakog NRG domena obezbeđuje razumevanje signalne
specifičnosti neuregulina. Kringle domeni, karakteristični samo za
tip II, sastoje se od trostruko uvijenih, 3-disulfidnih mostova,
koji su inače prisutni u faktorima koagulacije i smatra se da služe
kao protein-protein vezujuća polja. Imunoglobulinima slični domeni
imaju oko 80 aminokiselinskih rezidua formirajući 7–10 β podgrupa i
obezbeđuju različite ćelijske funkcije, uključujući molekularni
transport, adheziju, morfogenu kontrolu i identifikaciju ćelija.
Jedinstveni region svih neuregulina, EGF sličan domen, je proteinski
domen koji se sastoji od 6 cisteinskih rezidua, koje formiraju
3-disulfidne veze. Nrg-β izoforma je suštinski potentnija od Nrg-α
izoforme. Međutim to ne sugeriše da su Nrg-β izoforme biološki
irelevantnije, jer je potvrđeno da je Nrg-α neophodan u razvoju
dojki [7].
NRG-1 deluje na parakrini i jukstakrini način preko familije ErbB
tirozin kinaza receptora (ErbB2, ErbB3, i ErbB4). ErbB receptori
sadrže ekstracelularna mesta za ligande, transmembranske domene i
tirozin kinaza domene. NRG-1 vezujući se za ErbB3 i ErbB4 (ali ne i
za ErbB2), indukuje konformacione promene koje dozvoljavu
dimerizaciju ErbB2 ili aktivaciju ligandima ErbB3 ili ErbB4
receptora.
Značaj neuregulina u razvoju srca
Sredinom 90-ih godina ukazalo se na krucijalnu ulogu neuregulin/ErbB
signalnog puta u razvoju srca [8]. U prenatalnom srcu, NRG-1 i NRG-2
se sintetišu u endokardno endotelnim i mikrovaskularnim endotelnim
ćelijama u srcu. Nakon oslobađanja u ekstracelularni prostor,
neuregulin se vezuje za ErbB4 receptore prisutne na miokardu.
Vezivanje neuregulina-1 i -2 na ErbB4 promoviše proliferaciju i
diferencijaciju kardiomiocita u razvoju srca. Postoji jasno
razgraničenje u specifičnosti koja kontroliše ovu proliferaciju,
kako na nivou liganda, tako i na nivou receptora. Za prenatalni
razvoj srca neophodan je neuregulin-1 koji sadrži imunoglobulinima
slične domene. Kod knock out miševa evidentirano je da nedostatak
Ig-neuregulina-1 može biti odgovoran za nenormalno tanak miokradni
zid. Zanimljivo je da i pored ekspresije proteina NRG-2 u
prenatalnom srcu, to nije dovoljno za odvijanje kardiomiocitne
proliferaciju u odsustvu Ig-Nrg-1 proteina. Podatak ide u prilog
činjenici da nedostatak NRG-2 nije letalan za embrion niti ima
efekte na srce. Posmatrano na nivou receptora, nedostatak ErbB4 ili
ErbB2 rezultuje fenotipom koji je sličan nedostatku neuregulinu-1,
gde odsustvo bilo kojih receptora dovodi do nedovoljnog razvoja
miokardnog zida. Konačno, smanjenje debljine miokarda vidi se kod
nedostatka NRG-1, ErbB2 i ErbB4 što dovodi do insuficijencije
razvoja srca i trabekulacije zida komora. Nivo neuregulina-1 mora
biti jasno regulisan, jer ektopična sineteza neuregulina u razvoju
srca može dovesti do hipertrabekulacije i malformacija poznatih kao
nekompaktnost ventrikula, povezano sa brojnim kongenitalnim srčanim
bolestima. ErbB3 je detektovan u mezenhimu i srčanom endotelu tokom
razvoja srca, ali ne i kod odraslih. Ta posebna lokalizacija se
odražava na nenormalni razvoj srca kod miševa bez ErbB3. Pored
uticaja na proliferaciju i diferencijaciju kardiomiocita, NRG-1
takođe promoviše odgovarajuću diferencijaciju i regrutovanje ćelija
provodnog sistema srca od kontraktilnih kardiomiocitima [9].
Uloga neuregulina u fiziološkim i patofiziološkim procesima u
srcu
Neregulin-1 pripada epidermalnim faktorima rasta (EGF) koji imaju
ulogu u proliferaciji, diferencijacije i očuvanju funkcije epitelnih
ćelija dojke, glijalnih ćelija, neurona i miocita. Ovi biološki
efekti su posredovani preko seta tirozin kinaza ligand zavisnih
receptora (ErbB2, ErbB3, ErbB4) što dovodi do fosforilacije i
posledične signalizacije. Dobro je opisan signalni put NRG-1/ErbB
zbog neophodne uloge u razvoju srca i nervnog sistema. On ima svoje
mesto u patofiziologiji shizofrenije i nekih karcinoma. Prvi podaci
o značaju neuregulina kod ljudi dobijeni su nakon primene lekova
koji ostvaruju svoj efekat putem aktivacije ErbB2 receptora, takođe
poznat kao humani epidermalni faktor rasta-2/neu (HER-2). To su bili
pacijenti sa metastaskim tumorom karcinoma dojke koji su u terapiji
primali i monoklonska antitela anti ErbB2, lek trastuzumab
(herceptin), sam ili u kombinaciji sa antraciklinom. Kod ovih
bolesnika se češće javljala srčana insuficijencija i sistolna
disfunkcija komora [10]. Endogeni i egzogeni neuregulin-1 imaju
kardioprotektivne i kardioregenerativne funkcije i mogu da utiču na
apoptozu, nereguralnost miofibrila, antraciklinom indukovanu
kardiomiopatiju, formiranje ožiljka kao i proliferaciju kardimiocita
postnatalno (slika 1). Najvažniji efekti neuregulina u
kardioprotekciji i kardioregeneraciji su, u stvari, efekti faktora
rasta, poput efekta insulinu sličnog faktora rasta (IGF-1),
periostin peptidu, ili fibroblastični faktor rasta-1 (FGF-1).
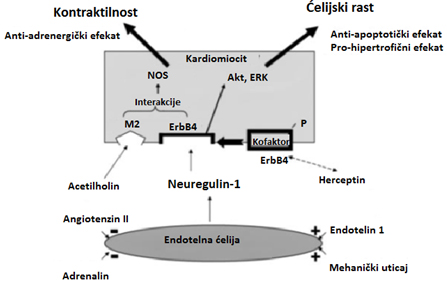
Slika 1. Mesto sinteze i uticaj neuregulina na srce.
NRG-1/ErbB sistem je odgovoran za fiziološku adaptaciju srca pri
promenjenim srčanim zahtevima. Ovaj koncept je objašnjen kroz
signalni put NRG-1/ErbB tokom adaptacije kardiovaskularnog sistema u
trudnoći. U eksperimentu na miševima pokazano je da se osovina
NRG/ErbB2 aktivira u trudnoći. Supresija ErbB sa ErbB tirozin kinaza
inhibitorima tokom trudnoće dovodila je do povećane ventrikularne
dilatacije, smanjenja sistolne funkcije i prevremenog uginuća
pregnantnih miševa. Ovim se potvrdila važnost intaktne
neuregulin-1/ErbB signalne osovine za adaptaciju srčane morgologije
i funkcije na nove uslove [11].
Takođe, in vitro studijama je objašnjen ćelijski mehanizam kojim
NRG/ErbB signalni put može regulisati adaptaciju srca na stres.
Zaključcima ove studije je potvrđen koncept da parakrini i
jukstakrini sistem regulišu interakcije između mikrovaskularnih
endotelnih ćelija i kardiomiocita. Brojne izoforme NRG-1 se
sintetišu u endotelnim ćelijama. Većinom je to tip I pre-neuregulina
čija se proteolitička aktivacija mora desiti u neuregulin-1 da bi se
iz endotelnih ćelija oslobodio aktivni ligand. Blagi oksidativni
stres izaziva oslobađanje i delovanje NRG-1, testirano u in vitro
uslovima, preko membranskih metaloproteaza, kao što je objašnjeno u
sistemu nervnih ćelija. Slično, drugi fiziološki stimulusi mogu da
utiču na oslobađanje i delovanje NRG-1, kao što su angiotenzin,
fenilefrin i endotelin [12]. Mada je izoforma NRG-1 izražena u
srčanim mikrovaskularnim endotelnim ćelijama, nije potpuno jasna
uloga koju ova forma neuregulina ima u srcu.
ErbB2 i ErbB4 receptori su distribuirani na kardiomiocitima adultnog
srca. ErbB3 receptori su takođe prisutni u adultnom miokardu, ali
njihova uloga nije poznata. Na izolovanim kardiomiocitima
rekombinantni neuregulin-1 β aktivira ErbB2 i ErbB4 receptore,
fosforilacija receptora dovodi do aktivacije ekstracelularnim
signalom regulisane kinaze (Erk), mitogenom aktivirane protein
kinaze (MAPK), fosfatidil-3 kinaze (PI3K), proteinske kinaze B (Akt)
i fokalne adhezion kinaze (FAK), dovodeći do promena u funkciji
miocita, uključujući ćelijsko preživljavanje, funkciju mitohondrija,
proliferaciju, rast, unos glukoze, nivo kalcijuma u sarkoplazmatskom
retikulumu i formiranje fokalnih pripoja. NRG-1β takođe reguliše
adrenergički odgovor kardiomiocita, ima negativan inotropni efekat
na izolovane papilarne mišiće i štiti miocite od ćelijske smrti
indukovane aktivacijom β1 receptora.
Poput neuregulina-1 i IGF-1, FGF-1, FGF-2, urocortin, VEGF, TGF
beta-1 i kardiotrofin-1 su povezani sa smanjenjem apoptoze u srcu.
Međutim, administracija Nrg-1 nakon ligature prednje leve koronarne
arterije nema efekta na apoptozu, dok kod tretmana dijabetične
kardiomiopatije u slučaju bivalente administracije NRG-1 nakon
antraciklinom indukovane kardiotoksičnosti izaziva smanjenje
kardiomicitne apoptoze. Kod pacijenata sa srčanom insuficijencijom,
nivo neuregulina je normalan, dok je ekstremno mali broj ErbB2 i
ErbB4. Ovi podaci sugerišu o značaju receptora u insuficijentnom
miokardu.
Endogeni neuregulin može takođe uticati na srčanu funkciju
delovanjem na nekardiomiocite. NRG-1 promoviše srčanu funkciju
utičući na angiogenezu. Endotelni neuregulin promoviše angiogenezu u
oba, u in vitro i in vivo uslovima, a smanjenje neuregulina u
angiogenezi je povezano sa ishemijskim oštećenjima, ukazujući na
ulogu neuregulina kao proangiogenetskog faktora. Neuregulin
stimuliše angiogenezu nezavisno od VEGF. Kardioprotektivni efekat
neuregulina-1 preko nekardiomiocita može se ostvariti povećanjem
aktivnosti srčanih fibroblasta, gde administracija neuregilina
rezultuje u smanjenju veličine ožiljka. Da li neuregulin deluje
direktno ili indirektno na fibroblaste nije poznato i predstavlja
još jedno polje za istraživanje kardioprotektivnog efekta NRG-1.
Kako neuregulin utiče na adultno srce ostaje intrigantno polje za
istraživače. Endotelin-1 povećava kontraktilnost srca i povećava
ekspresiju neuregulina, dok je angiotenzin II i fenilefrin smanjuju
[13]. Ovo govori o važnosti mehanizma za regulaciju neuregulin
signalnog puta nivoom liganda koji određuje veličinu srčanih
zahteva. Integirni su familija receptora na mnogim ćelijama, pa i na
kardiomiocitima, koji su osetljivi na mehanički stres i mogu biti
netipični receptori za NRG-1. Ekspresija neuregulina u trudnoći je
povećana u vreme kada hemodinamski stres povećava srčane zahteve.
Paralele su viđene između neuregulina i drugih faktora rasta
potvrđujući protektivnu ulogu na srce, što može biti značajno u
rasvetljavanju terapeutskog potencijala srčanih faktora rasta (slika
2).
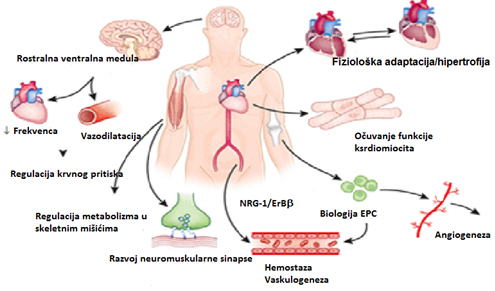
Slika 2. Fiziološka i patofiziološka uloga neuregulina-1 na
kardiovaskularni system.
Endotelne progenitorne ćelije (EPC) poreklom iz koštane srži
igraju ulogu u obnavljanju tkiva nakon ishemijskog oštećenja.
Ishemijska oštećenja kardiomiocita i fizičko opterećenje dovode do
aktivacije tkivnog neuregulina-1 i potentni su induceri mobilizacije
EPC i regrutovanja u srčane i skeletne mišiće [14]. Interesantno je
da endotelne progenitorne ćelije imaju ErbB2 i ErbB3 receptore, koje
aktivira NRG-1. U kojoj meri NRG-1 modulira funkciju endotelnih
ćelija u stanjima kao što je oporavak miokarda nakon ishemije ostaje
da se potvrdi u narednim istraživanjima.
Uloga neuregulina kao potencijalnog biomarkera
Poznati su značaj i uloga neuregulina u skeletnim mišićima tokom
opterećenja, u srcu tokom oštećenja, kao i pri promeni hemodinamskog
opterećenja. Ovo navodi na činjenicu da bi bilo poželjno izmeriti
nivo neuregulina-1 u plazmi ili serumu. U zdravih osoba, nivo
neuregulina-1 je povezan sa fizičkom utreniranošću i stanjem
zdravlja. Tako, nivo neuregulina korelira sa prognozom bolesti kod
osoba sa srčanom insuficijencijom. Takođe, viši nivo neuregulina
povezan je sa lošijom prognozom ishemijske bolesti srca. U
pacijenata sa stabilnom koronarnom bolešću bez ishemije ili srčane
insuficijencije, nivo neuregulina-1 obrnuto je povezan sa težinom
koronarne arterijske bolesti. Interesantno je da je nivo
neuregulina-1 bio viši u pacijenata sa stres indukovanom ishemijom
(15). Moguće je da nivo neuregulina u plazmi može biti klinički
indikator u bolestima srca i kardiovaskularnog sistema.
Neuregulin u terapiji kardiovaskularnih bolesti
S obzirom na to da je neuregulin neophodan u razvoju
kardiovaskularnog sistema, regulaciji kardioprotektivnih signalnih
mehanizama, moguće je da aplikacija rekombinantnih neuregulin
izoformi može imati potencijalnu primenu u terapiji pojedinih
srčanih oboljenja (slika 3).
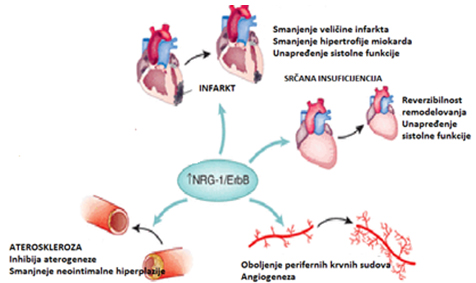
Slika 3. Potencijalni efekti neuregulina-1 u terapiji
kardiovaskularnih bolesti.
Ima više kliničkih ispitivanja o administraciji NRG-1 kao srčanoj
terapiji, ali je uglavnom on ograničen na lečenje sistolne
disfunkcije. Eksperimenti na životinjama sa dijabetičnom
kardiomiopatijom i angiotenzin-II izazvanim oštećenjem pokazali su
da administracija NRG-1 može da ublaži dijastolnu disfunkciju.
Imajući u vidu visoku učestalost dijastolne disfunkcije, buduća
istraživanjau kojima NRG-1 tretman može ublažiti disfunkcija
predstavlja važnu oblast istraživanja. Treba istražiti primenu
neuregulina u terapiji dilatirane i hipertofične kardiomiopatije. Da
li tretman NRG-1 može da poboljšaju restriktivnu i aritmogenu
kardiomiopatiju je nepoznato, i predstavlja još jedan intrigantan
prostor za buduće studije. Poznato je da neuregulin povećava broj
kardiomiocita, a ne njihovu veličinu, ali tačan mehanizam
poboljšanja srčane funkcije neuregulinom u akutnoj srčanoj
insuficijenciji nije poznat [16].
Kritičko pitanje za primenu terapije neuregulinom je kako, kada i u
kojim bolestima ga primeniti? Najveći benefit terapije neuregulinom
na srce je nakon oštećenja miokarda. Neuregulin pormoviše
preživljavanje kardiomiocita. U terapiji karcinoma antraciklinom,
koji je kardiotoksičan i čija administracija oštećuje srce,
neuregulin ima kardioprotektivni efekat na srce. Otuda se sugeriše
koterapija antraciklina i neuregulina [17].
Međutim primena rekombinantnog NRG-1 može da stimuliše proliferaciju
i nesrčanih ćelija. Pitanje je i da li će upotreba neuregulina u
kardioprotekciji aktivirati samo za to namenjene receptore jer ako
se NRG-1 daje sistemski može da aktivira receptore u drugim tkivima
i promoviše neželjene reakcije [18].
Genetska arhitektura neuregulina omogućava postojanje brojnih
izoformi. Različite izoforme imaju i preklapajuće i drugačije
efekete, što odražava kompleksnost Nrg/ErbB signalnog puta.
Različite efekti neuregulina koji mogu da budu iskoristivi za
kliničku primenu sa višestrukim efektima na srce pokazuju velike
mogućnosti za lečenje srčane insuficijencije.
LITERATURA
- Parodi EM, Kuhn B. Signalling between microvascular
endothelium and cardiomyocytes through neuregulin. Cardiovasc
Res, 2014; 102 (2): 194–204, doi: 10.1093/cvr/cvu021.
- Hertig CM, Kubalak SW, Wang Y, Chien KR. Synergistic roles
of neuregulin-1 and insulin-like growth factor-I in activation
of the phosphatidylinositol 3-kinase pathway and cardiac chamber
morphogenesis. J Biol Chem, 1999; 274: 37362–37369.
- Lee KF, Simon H, Chen H, Bates B, Hung MC, Hauser Cetal.
Requirement for neuregulin receptor erbB2 in neuraland cardiac
development. Nature, 1995; 378: 394–398. 68.
- Gassmann M, Casagranda F, Orioli D, Simon H, Lai C, Klein R
et al. Aberrant neural and cardiac development in mice lacking
the ErbB4 neuregulin receptor. Nature, 1995; 378: 390–394.
- Marchionni MA, Goodearl AD, Chen MS, Bermingham-McDonogh O,
Kirk C, Hendricks M et al. Glial growth factors are
alternatively spliced erbB2 ligands expressed in the
nervoussystem. Nature 1993;362:312–318.
- Steinthorsdottir V, Stefansson H, Ghosh S, Birgisdottir B,
Bjornsdottir S, Fasquel AC et al. Multiple novel transcription
initiation sites for NRG1. Gene 2004;342:97–105.
- Li L, Cleary S, Mandarano MA, Long W, Birchmeier C, Jones F
Eetal. The breast protooncogene, HRG alpha regulates epithelial
proliferation and lobulo alveolar development in the mouse
mammary gland. Oncogene 2002;21:4900–4907.
- Lee KF, Simon H, Chen H, Bates B, Hung MC, Hauser C.
Requirement for neuregulin receptor erbB2 in neural and cardiac
development. Nature 1995; 378(6555):394–8.
- Emily M. Parodi ME, Kuhn B. Signalling between microvascular
endothelium and cardiomyocytes through neuregulin.
Cardiovascular Research 2014 ;102:194–204 doi:10.1093/cvr/cvu021
- Slamon DJ, Leyland-Jones B, Shak S, Fuchs H, Paton V,
Bajamonde A, Fleming T, Eiermann W, Wolter J, Pegram M, Baselga
J, Norton L. Use of chemotherapy plus a monoclonal antibody
against HER2 for metastatic breast cancer that overexpresses
HER2. N Engl J Med 2001; 344:783–792. [PubMed: 11248153]
- Lemmens K, Doggen K, De Keulenaer GW. Activation of the
neuregulin/ErbB system during physiological ventricular
remodeling in pregnancy. Am J Physiol Heart Circ Physiol, 2011;
300: H931–H942. [PubMed: 21186272].
- Lemmens K, Segers VF, Demolder M, De Keulenaer GW. Role of
neuregulin-1/ErbB2 signaling in endothelium-cardiomyocyte
cross-talk. J Biol Chem, 2006; 281: 19469–19477. [PubMed:
16698793].
- Smiljic S, Radović D, Nestorović V, Milanović Z, Biševac B.
Endotelini kao medijatori u modulaciji srčanih performansi.
Praxis medica, 2014; 3 (4): 79–84.
- L ee PSS, Poh KK. Endothelial progenitor cells in
cardiovascular diseases. World J Stem Cells, 2014. July 26; 6
(3): 355–366, doi: 10.4252/wjsc.v6.i3.355
- Rupert CE, Coulombe KL. The Roles of Neuregulin-1 in Cardiac
Development, Homeostasis, and Disease. Biomark Insights. 2015;
10 (1): 1–9. doi: 10.4137/BMI.S20061.
- Lemmens K, Doggen K, De Keulenaer GW. Role of
neuregulin-1/ErbB signaling in cardiovascular physiology and
disease: implications for therapy of heart failure. 2007; 116
(8): 954–60.
- Noireaud J, Andriantsitohaina R. Recent Insights in the
Paracrine Modulation of Cardiomyocyte Contractility by Cardiac
Endothelial Cells. BioMed Research International, 2014, Article
ID 923805, 10 pages
http://dx.doi.org/10.1155/2014/923805
- Odiete O, Hill MF, Sawyer DB. Neuregulin in cardiovascular
development and disease. Circ Res, 2012; 111 (10): 1376-85, doi:
10.1161
|
|
|
|
